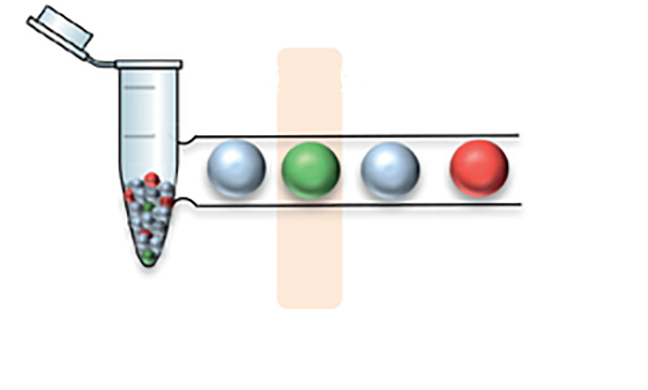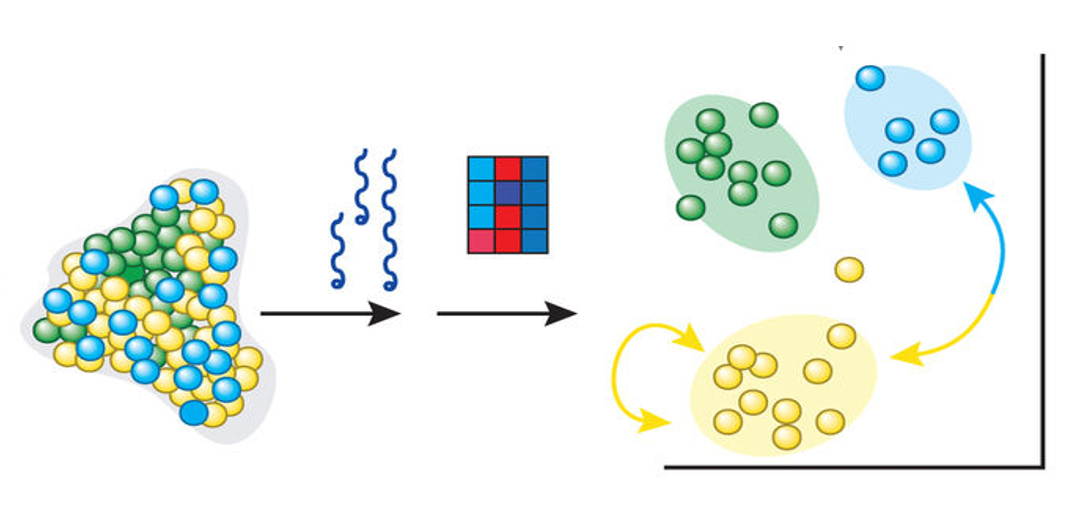
Számos molekuláris genetikai diagnosztika első lépése a biológiai mintából történő DNS izolálása. Genetikai tanácsadás során, amikor öröklődő megbetegedés gyanúja merül fel, az esetek nagy részében a sejtmagban elhelyezkedő DNS (ritkábban mitokondriális DNS) szekvenciájában vagy metilációs mintázatában keressük a referenciagenomhoz hasonlítva a vizsgálandó DNS eltéréseit. Az egyes biológiai mintákból történő DNS izolálás (vagy extrakció) kivitelezéséhez számos különböző gyártó által forgalmazott kit van forgalomban, amelyek gyors és megbízható, akár automatizált rendszerek alkalmazásával, DNS izolálást tesznek lehetővé.
A DNS izolálás fő lépései: 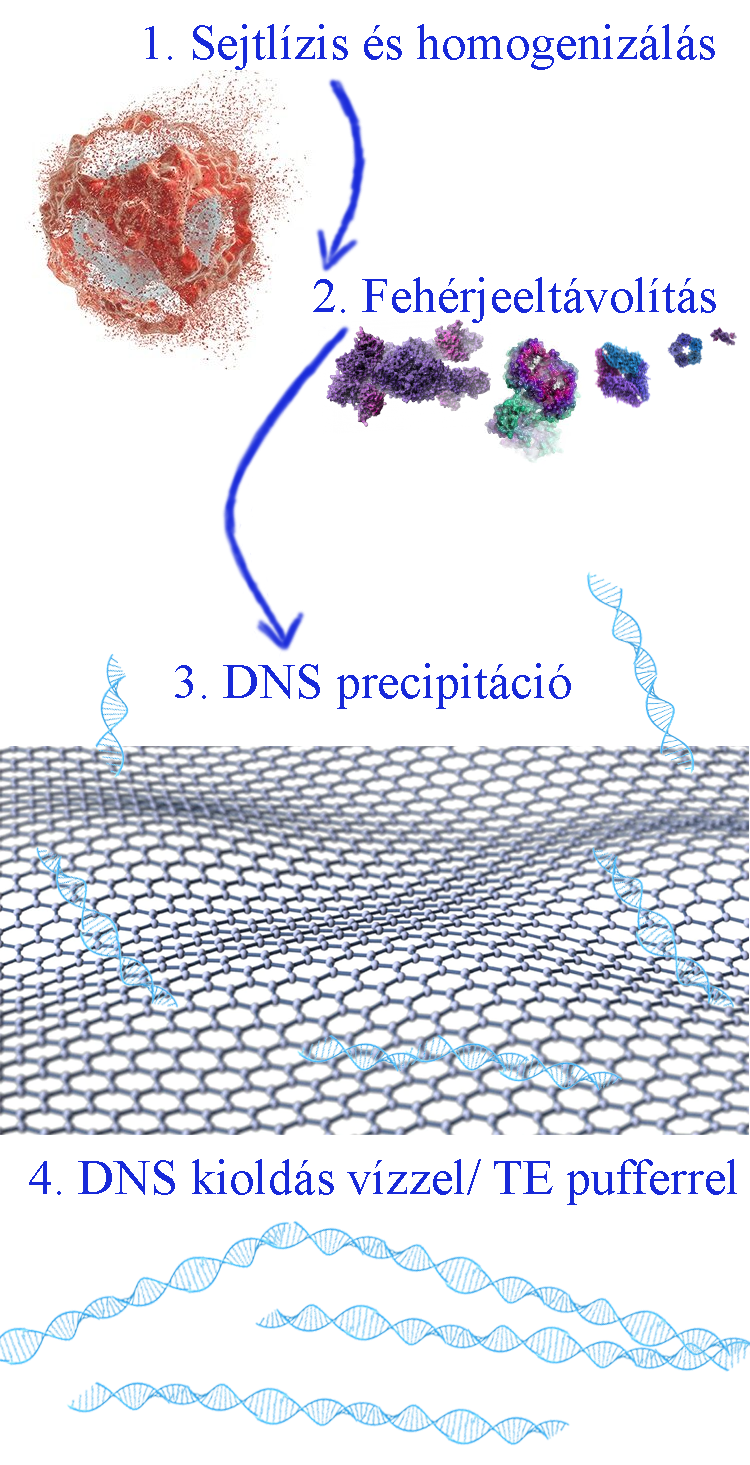 Kiemelendő, hogy a DNS izolálása után fontos az izolált DNS pontos mennyiségének, tisztaságának és minőségének az ellenőrzése, mivel számos módszer előfeltétele az izolált DNS minta épsége, amely nem tartalmaz RNS vagy fehérje szennyezést, valamint az izolálás során alkalmazott reagens, oldat kontamináció mentes (pl. alkohol-szennyezés). Az izolálás mennyiségi és minőségi meghatározására: (1) kétszálú DNS-re specifikus (dsDNS) festék alkalmazásával fluoriméterrel határozzuk meg a koncentrációt, (2) a tisztaságot és a kontamináció kizárását spektrofotométerrel az A260/A280, illetve A260/A230 hullámhosszon történő elemzéssel vizsgáljuk, (3) valamint a DNS agar-agar gélen történő megfuttatása DNS létra jelenlétében a fragmentáció kizárást teszi lehetővé.
Kiemelendő, hogy a DNS izolálása után fontos az izolált DNS pontos mennyiségének, tisztaságának és minőségének az ellenőrzése, mivel számos módszer előfeltétele az izolált DNS minta épsége, amely nem tartalmaz RNS vagy fehérje szennyezést, valamint az izolálás során alkalmazott reagens, oldat kontamináció mentes (pl. alkohol-szennyezés). Az izolálás mennyiségi és minőségi meghatározására: (1) kétszálú DNS-re specifikus (dsDNS) festék alkalmazásával fluoriméterrel határozzuk meg a koncentrációt, (2) a tisztaságot és a kontamináció kizárását spektrofotométerrel az A260/A280, illetve A260/A230 hullámhosszon történő elemzéssel vizsgáljuk, (3) valamint a DNS agar-agar gélen történő megfuttatása DNS létra jelenlétében a fragmentáció kizárást teszi lehetővé.
RNS izolálás és reverz transzkripció cDNS-sé
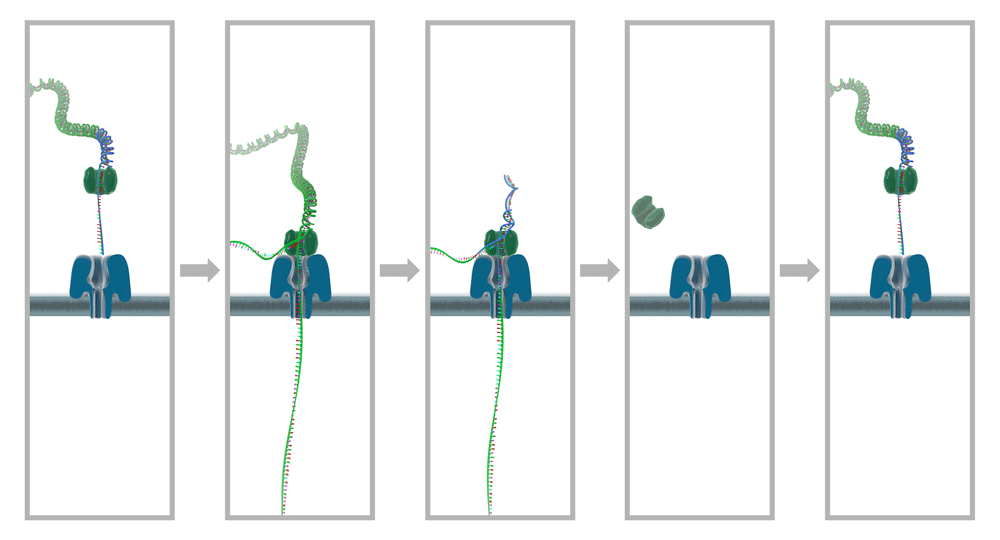
A humán sejtekben az mRNS-t, miRNS-t és más RNS-t kódoló gének sejttípus- és időfüggő módon fejeződnek ki, amelynek első nagy lépése a kérdéses gének transzkripciója (átírása). A transzkripció a sejtmagban történik, majd a folyamat végén az érett RNS molekulák a citoplazmába kerülnek. Az in vitro molekuláris vizsgálatok során a teljes RNS vagy bizonyos RNS típusok izolálása biofolyadékból (pl. vérplazma, likvor, vizelet) vagy különböző módszerekkel szeparált sejtek lízisét követően végezhető el. A mintafeldolgozás időzítése, kulcsfontosságú, mivel a különböző sejtstresszt kiváltó tényezőkre a döntően egyszálú molekulákat tartalmazó RNS mintázat nagy mértékben megváltozhat vagy károsodhat relatív rövid időn belül. Ezért igen kitünetett fontosságú az izolált RNS mennyiségi és minőségi ellenőrzése: (1) az RNS vagy RNS altípus specifikus festék alkalmazásával határozzuk meg az adott RNS koncentrációját fluoriméterrel, (2) a tisztaságot és a kontamináció kizárását spektrofotométerrel az A260/A280, illetve A260/A230 hullámhosszon történő elemzéssel vizsgáljuk, (3) valamint az RNS agar-agar gélen történő megfuttatása molekulasúly marker létra jelenlétében az izolált RNS molekulák méreteinek meghatározása javasolt.
Az izolált RNS olykor vizsgálható natívan is, például nanopórus szekvenálással (amely feltárja a mintában található RNS molekulák szekvenciáit és ezek epigenetikai módosításait), de az RNS-t vizsgáló molekuláris biológiai módszerek legtöbbje az RNS-ből átírt cDNS molekulákat vizsgálja. 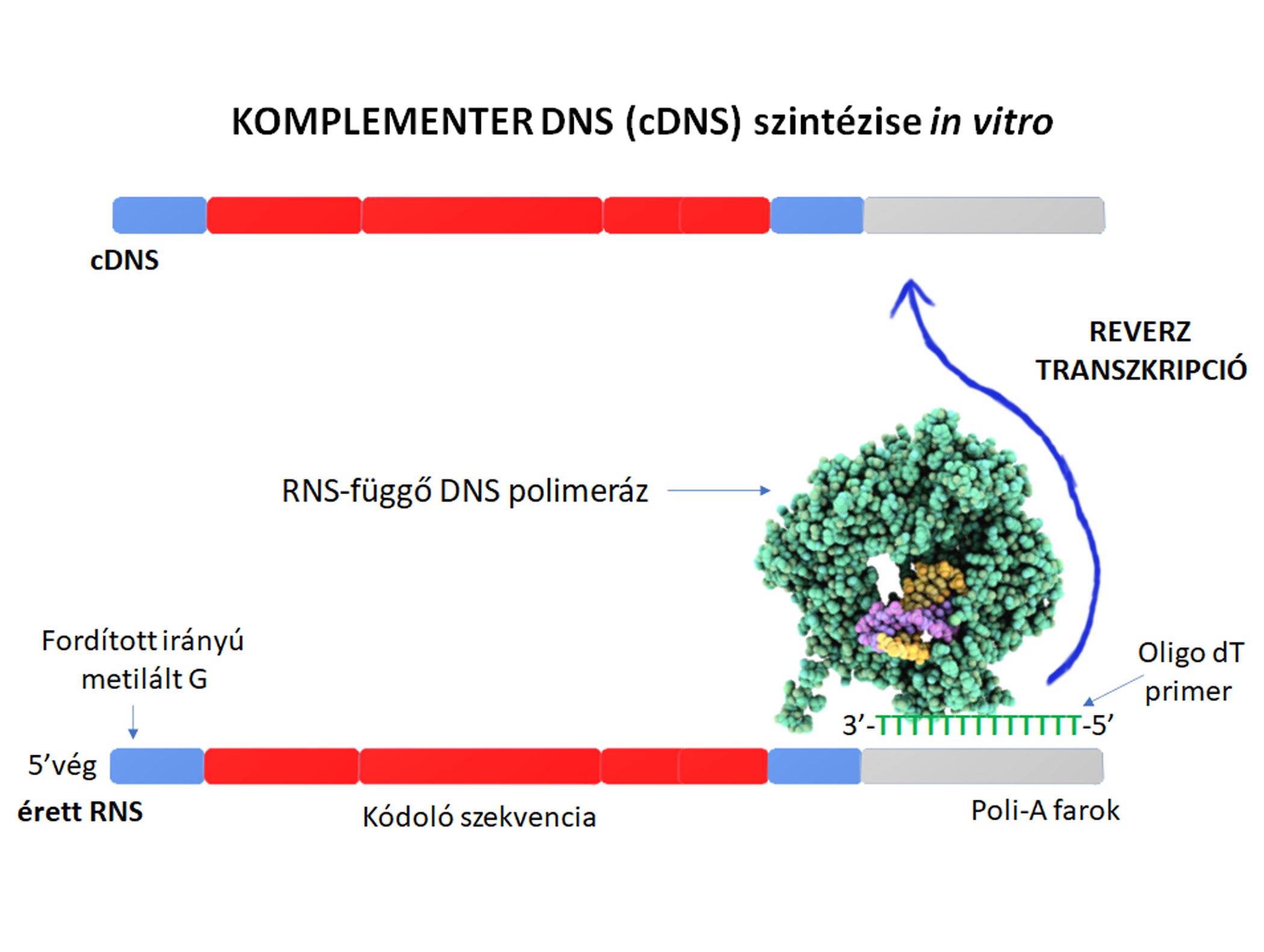 A reverz transzkripció során képződött cDNS molekulák stabilak, hosszú távon tárolhatók 4 és -20 Celsius fok között. Endogén vagy exogén kontrollok alkalmazásával meghatározható egy adott RNS relatív vagy abszolút szintje, amelyből bizonyos esetekeben diagnosztikus értékű lehet, vagy további génvizsgálatok elvégzésére hívhatja fel a szakember figyelmét. Szekvenálással pedig meghatározható a mintában található RNS mintázat.
A reverz transzkripció során képződött cDNS molekulák stabilak, hosszú távon tárolhatók 4 és -20 Celsius fok között. Endogén vagy exogén kontrollok alkalmazásával meghatározható egy adott RNS relatív vagy abszolút szintje, amelyből bizonyos esetekeben diagnosztikus értékű lehet, vagy további génvizsgálatok elvégzésére hívhatja fel a szakember figyelmét. Szekvenálással pedig meghatározható a mintában található RNS mintázat.
1993-ban a kémiai Nobel-díjat a PCR módszer kidolgozásáért adták. Molekuláris genetikai alaptechnika, amely során egy adott cél DNS molekularészlet milliószoros nagyságrendű felsokszorozása (amplifikációja) valósul meg. Az amplifikálható DNS szakasz hossza 50 - 10 000 bp között mozoghat. A módszer 3 fő lépés 35-45-szörös (lehet ennél kisebb számú ciklusokból álló PCR is, pl. szekvenálási könyvtárkészítés során) ismétléséből áll: (1) denaturáció (a kétszálú templát DNS magas, azaz 94-96 Celsius fokon, történő hevítése, amelynek eredményeként a DNS molekula két szála kettéválik), (2) primer kapcsolódás (45-60 Celsius fokon az egyszálú DNS-hez a primerek a komplementaritás alapján hibridizálnak), és (3) extenzió (72 Celsius fokon a hőrezisztens DNS függő DNS polimeráz, például a Taq polimeráz, a primerektől elindulva a DNS szál szintézisét elvégzi). 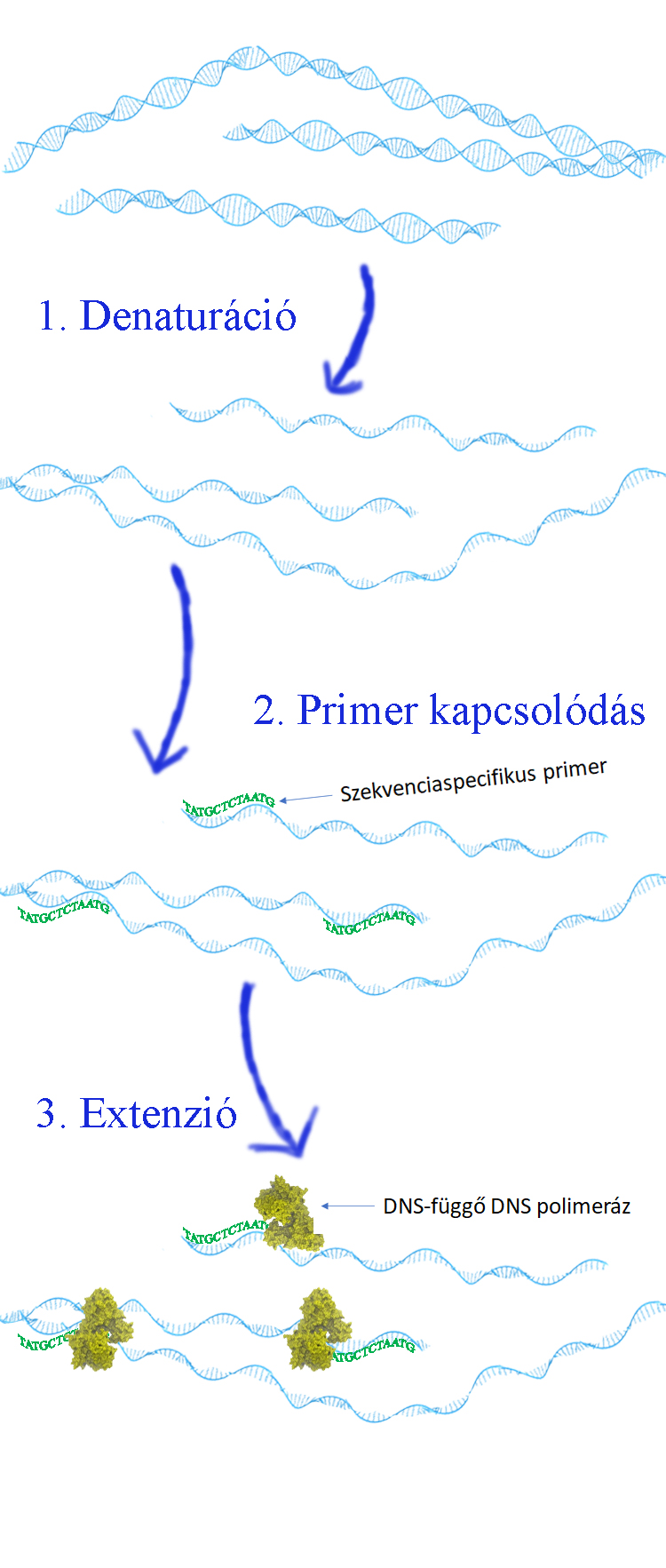 A kapott PCR terméket elemezhetjük, különböző restrikciós endonukleázokkal (pontmutáció elemzés), tovább tisztíthatjuk majd megszekvenálhatjuk (szekvencia analízis), meghatározhatjuk az abszolút mennyiséget (például telomérhossz; adott gének kópiaszámát- amely fontos lehet bizonyos betegség súlyosságának, várható progresszíójának megbecsléséhez vagy rizikó meghatározáshoz) vagy más molekuláris biológiai módszerrel vizsgálhatjuk a PCR termékeket.
A kapott PCR terméket elemezhetjük, különböző restrikciós endonukleázokkal (pontmutáció elemzés), tovább tisztíthatjuk majd megszekvenálhatjuk (szekvencia analízis), meghatározhatjuk az abszolút mennyiséget (például telomérhossz; adott gének kópiaszámát- amely fontos lehet bizonyos betegség súlyosságának, várható progresszíójának megbecsléséhez vagy rizikó meghatározáshoz) vagy más molekuláris biológiai módszerrel vizsgálhatjuk a PCR termékeket.
Fluoreszcens in situ hibridizáció (FISH)
Fluoreszcens mikroszkóp segítségével láthatóvá tehetők adott cél DNS molekularészletek vagy bizonyos cél RNS molekulák, valamint meghatározható in situ a megcélzott nukleinsavszekvenciák kópiaszáma és intracelluláris lokalizációja. Ezt a módszert elsődlegesen kromoszómaszámszám vagy a kromoszómák szerkezeti rendellenségek (pl. 21-es, 13-as vagy 18-as triszómia, X monoszómia, részleges 5. kromoszóma rövid kar monoszómia, különböző kromoszómák közötti átrendeződések vagy egy adott kromoszómán belüli rendellenes szerkezeti eltérések) gyanúja esetén alkalmazzák. A módszer első lépéseként a vizsgálni kívánt sejteket centrifugáljuk, majd egy tárgylemezre cseppentjük. 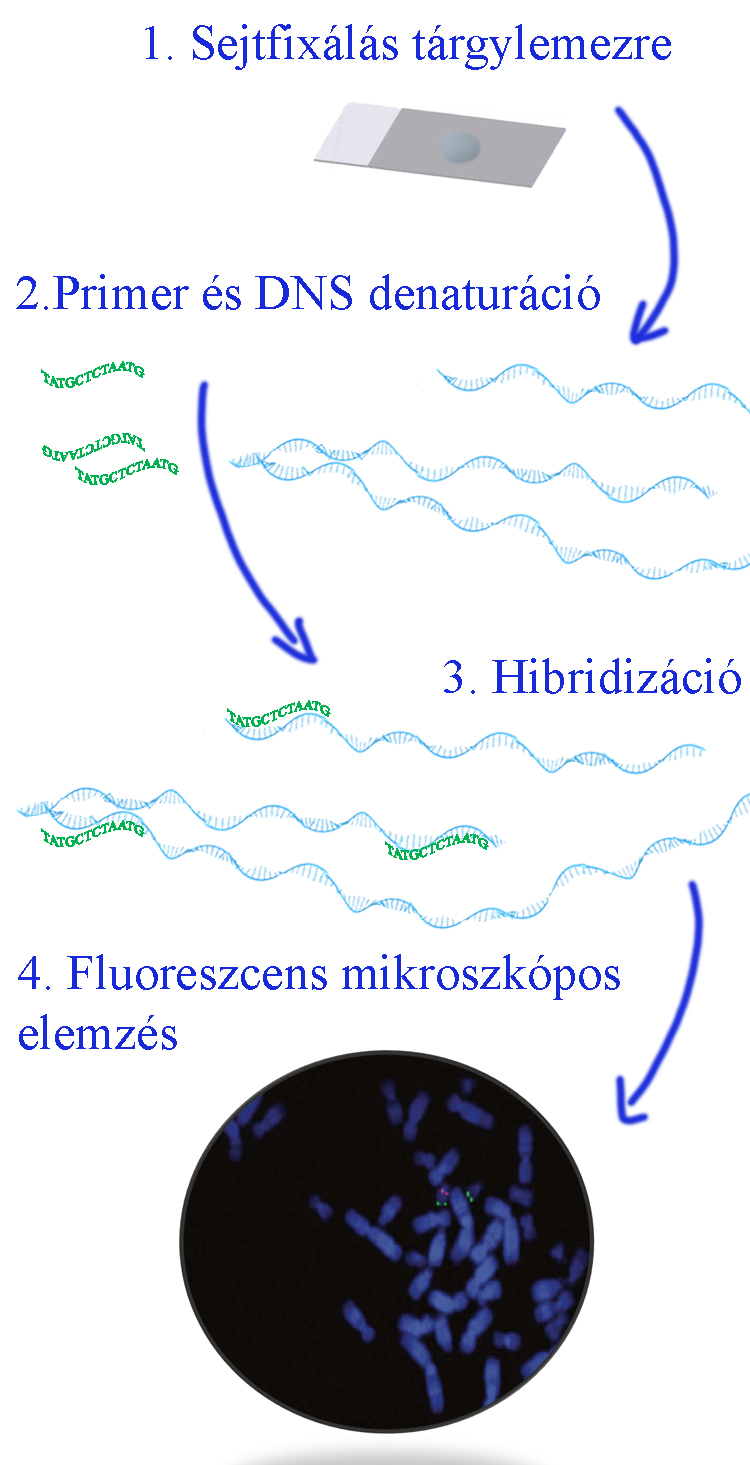 Második lépésben megcélzott DNS szekvenciára specifikus fluorokrómmal jelölt primer próbát és a vizsgálandó sejtek DNS molekuláit denaturáljuk (a két szálú molekulákat szétválasztjuk), majd a következő lépésben a primer próbákat a cél DNS molekulákkal hibridizáljuk és fluoreszcens mikroszkóppal vizsgáljuk.
Második lépésben megcélzott DNS szekvenciára specifikus fluorokrómmal jelölt primer próbát és a vizsgálandó sejtek DNS molekuláit denaturáljuk (a két szálú molekulákat szétválasztjuk), majd a következő lépésben a primer próbákat a cél DNS molekulákkal hibridizáljuk és fluoreszcens mikroszkóppal vizsgáljuk.

Komparatív genom hibridizáció (CGH)
A CGH technika lényege ugyanazon körülmények között egy adott mintából izolált DNS-sel (vagy cDNS) párhuzamosan számos hibrizációs esszé futtatása. A módszer alkalmas körülbelül 1 Mb-nál nagyobb DNS szakaszok deléciójának, duplikációjának vagy amplifikációjának, illetve egyidejűleg több ezer egy nukelotid polimorfizmusok/ variációk és a sejtek RNS expressziójának a meghatározására.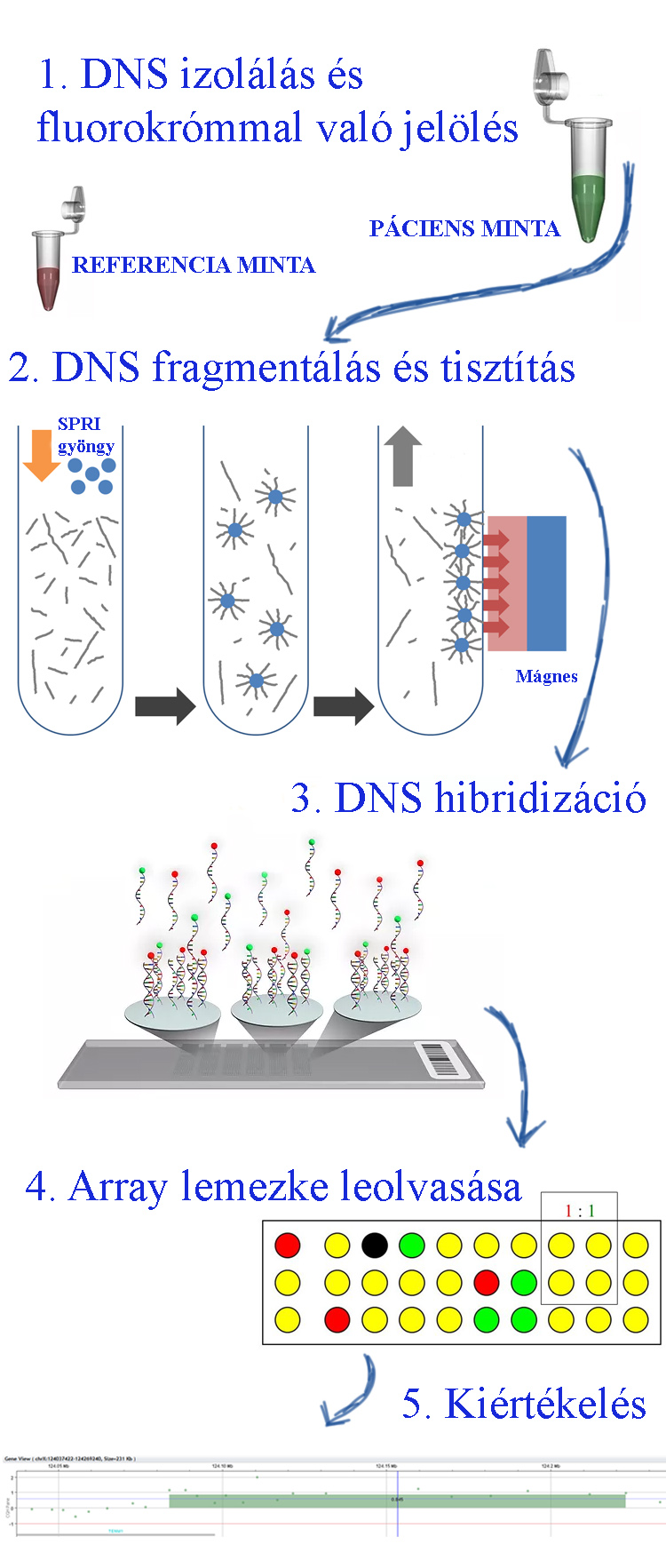 A mintából DNS-t izolálunk, a DNS-t denaturáljuk, majd flurokróm festékkel (pl. cianin-3) megjelöljük. Második lépésben a DNS molekulákat fragmentáljuk és SPRI gyöngyökhöz kötjük a kívánt méretű DNS szakaszokat. Centrifugálási lépésekkel tisztítjuk az SRPI gyöngyökhöz kötött DNS molekulákat, végül a gyöngyökről eluáljuk a DNS-t. A harmadik lépésben az array lapkán rögzítettek specifikus oligonukleotidokhoz hibridzáljuk a DNS molekulákat, majd 4. lépésben lézer szkennerrel leolvassuk és ezt követően értelmezzük a kapott adatokat.
A mintából DNS-t izolálunk, a DNS-t denaturáljuk, majd flurokróm festékkel (pl. cianin-3) megjelöljük. Második lépésben a DNS molekulákat fragmentáljuk és SPRI gyöngyökhöz kötjük a kívánt méretű DNS szakaszokat. Centrifugálási lépésekkel tisztítjuk az SRPI gyöngyökhöz kötött DNS molekulákat, végül a gyöngyökről eluáljuk a DNS-t. A harmadik lépésben az array lapkán rögzítettek specifikus oligonukleotidokhoz hibridzáljuk a DNS molekulákat, majd 4. lépésben lézer szkennerrel leolvassuk és ezt követően értelmezzük a kapott adatokat.
Bionano genomikai optikai térképezés
Strukturális variánsoknak tekintjük az 1000 bázispárnál nagyobb méretű DNS szekvencia eltéréseket a referencia genomtól. Az eltérés természete lehet mennyiségi (DNS szekvencia többlet pl. duplikáció, amplifikáció vagy hiány pl. deléció) vagy minőségi (DNS szekvencia kóros sorrend pl. inverzió, transzlokáció).Az új generációs szekvenálás vagy komparatív genomhibridizációval nem detekátált strukturális variánsok (pl. kiegyensúlyozott transzlokációk) azonosítására alkalmas módszer. Ezen strukturális variánsok optikai térképzése során a vizsgálandó mintából DNS-t izolálnak, majd a DNS molekulákat üveg felszínre rögzítik és kifeszítik nanocsatornákon.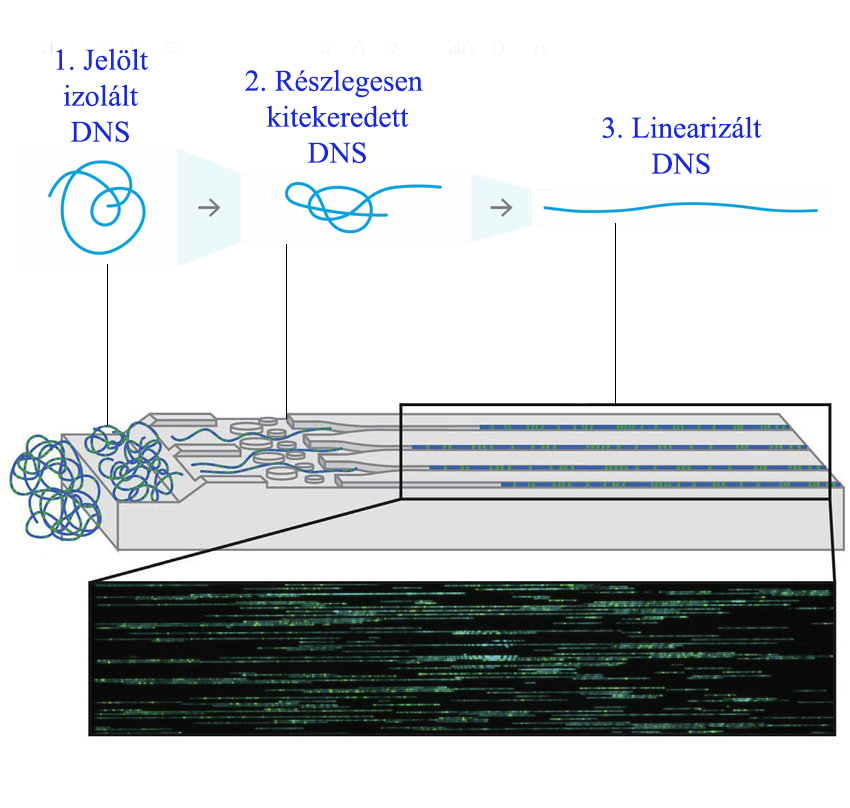 Ezen módszerrel jól vizsgálhatók a kromoszómák nehezen vizsgálható például a szubtelomérikus régiók. A módszer első lépése a DNS izolálásból áll, amely a gyártó által szolgáltatott kittel történik paramágneses lemez alkalmazásával (így elkerülhető a klasszikus centrifugálással történő szilika-alapú oszlopos módszer során keletkező részleges DNS fragmentáció). Ezt követő lépésben a DNS-t jelöljük specifikus szekvencia motívumoknál, amely nélküzhetetlen a megfelelő szkenneléshez és szekvencia-azonosításhoz. Következő lépésben az áramlási cellába pipettázva a DNS-t megtörténik a molekula linearizációja. A feltekeredett DNS először a mikrocsatornákba kerül, ahol részlegesen kitekeredik, majd a nanocsatornákba kerülve linearizálttá válik. Végül a jelölt linearizált DNS molekulák párhuzamosan szkennelésre kerülnek, majd motívum specifikusan a szekvencia mintázat azonosításea kerül bioinformatikai eszközök segítségével.
Ezen módszerrel jól vizsgálhatók a kromoszómák nehezen vizsgálható például a szubtelomérikus régiók. A módszer első lépése a DNS izolálásból áll, amely a gyártó által szolgáltatott kittel történik paramágneses lemez alkalmazásával (így elkerülhető a klasszikus centrifugálással történő szilika-alapú oszlopos módszer során keletkező részleges DNS fragmentáció). Ezt követő lépésben a DNS-t jelöljük specifikus szekvencia motívumoknál, amely nélküzhetetlen a megfelelő szkenneléshez és szekvencia-azonosításhoz. Következő lépésben az áramlási cellába pipettázva a DNS-t megtörténik a molekula linearizációja. A feltekeredett DNS először a mikrocsatornákba kerül, ahol részlegesen kitekeredik, majd a nanocsatornákba kerülve linearizálttá válik. Végül a jelölt linearizált DNS molekulák párhuzamosan szkennelésre kerülnek, majd motívum specifikusan a szekvencia mintázat azonosításea kerül bioinformatikai eszközök segítségével.
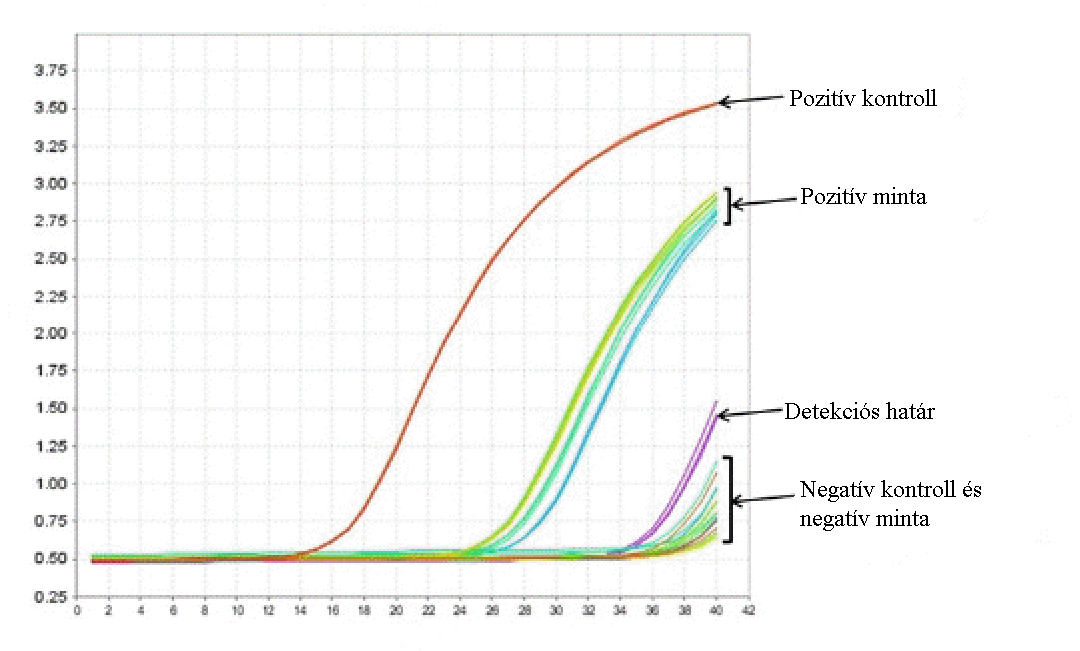
Kvantitatív polimeráz láncreakció (qPCR)
A valós idejű PCR igen széleskörűen alkalmazott technika.
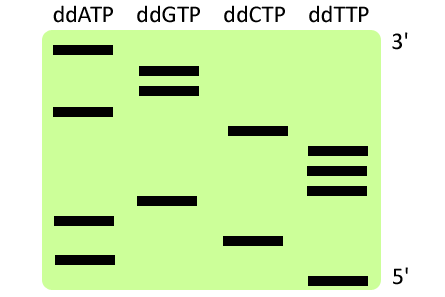
Sanger szekvenálás (I. generáció)
1980-ban a kémiai Nobel-díjat megosztva a Sanger szekvenálás kidolgozásáért adták.
CRISPR-Cas9 alapú genomszerkesztés
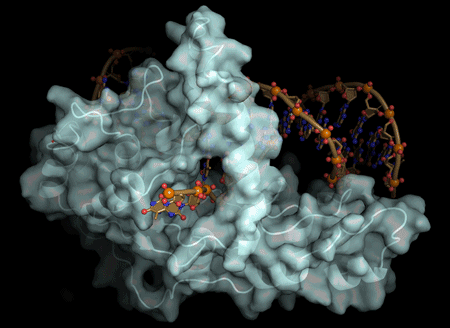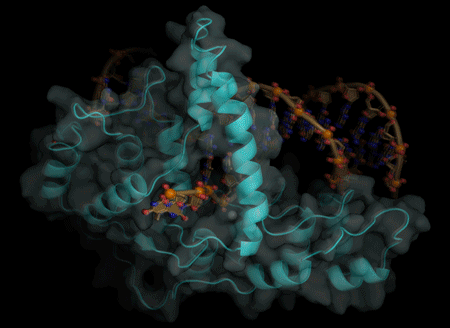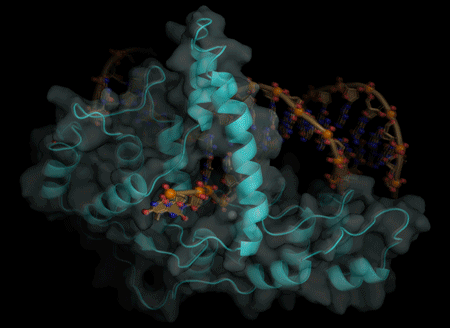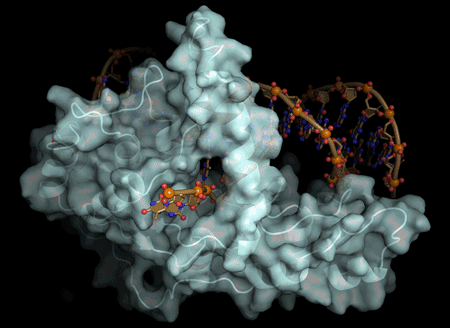
2020-ban a kémiai Nobel-díjat a CRISPR technológia kidolgozásáért adták. A CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) Cas9 rendszer 2012 óta alkalmazható génsebészeti eljárás, amely segítségével precíz célzott változást lehet létrehozni a sejtek nukleáris genomjába. A rendszer egyedi előnye az ún. multiplex genomsebészet lehetősége, amellyel egy időben több célgén is megváltoztatható. A rendszer alkalmazása óta folyamatos fejlesztéseket dolgoznak ki egy még hatékonyabb génszerkesztés érdekében: 1. Olyan Cas9 variánsok alkalmazása, amelyek csökkentik az ún. off target hatást (precizitást növelő javítás) 2. Egy adott gén transzkripciós aktiválása (CRISPRa) 3. Egy adott gén transzkripciós csendesítése (CRISPRi). A CRISPR-Cas9 rendszer segítségével inaktiválhatunk (kikapcsolhatunk) géneket, új gén(változatokat) adhatunk a genomhoz vagy génexpressziót fokozhatunk (bekapcsolhatunk) amely potenciálisan az embert érintő számos betegség végleges gyógyításának igéretét hordozza magában.

Adeno-asszociált vírus (AAV) molekuláris terápia
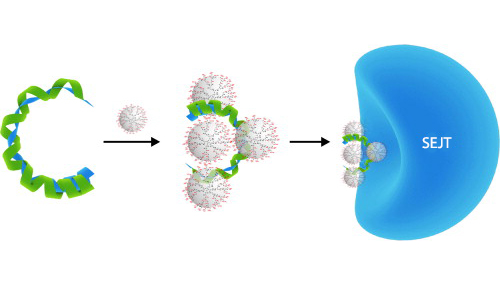
Az AAV-k a legkisebb vírusok közé tartoznak, átmérőjük mindössze 20 nm. Az AAV-k fehérjeburka ikozahedrális szerkezetű, a vírus belsejében pedig egyszálú DNS található, amely csak 4,7 Kb hosszú. Fontos megkülönböztetni az AAV-ket az adenovírusoktól. Az AAV immunogenitása gyenge, tehát nem vált ki erős immunválaszt, további előnyös tulajdonságai mellett képes egyaránt osztódó és nem osztódó sejtekbe bejutni. A bejutást követően a sejtmagban képes a nukleáris genomba integrálódni a genomba vagy a nukleáris genom mellett ún. episzómálisan maradni. A terápiásan bejutatott AAV vektor segítségével bejutatott génpótlás hosszan tartó génkifejeződést eredményez. Az AAV-k többféle szerotípusa ismert, melyekkel különböző (terápiásan kívánt) szöveteket lehet megcélozni.
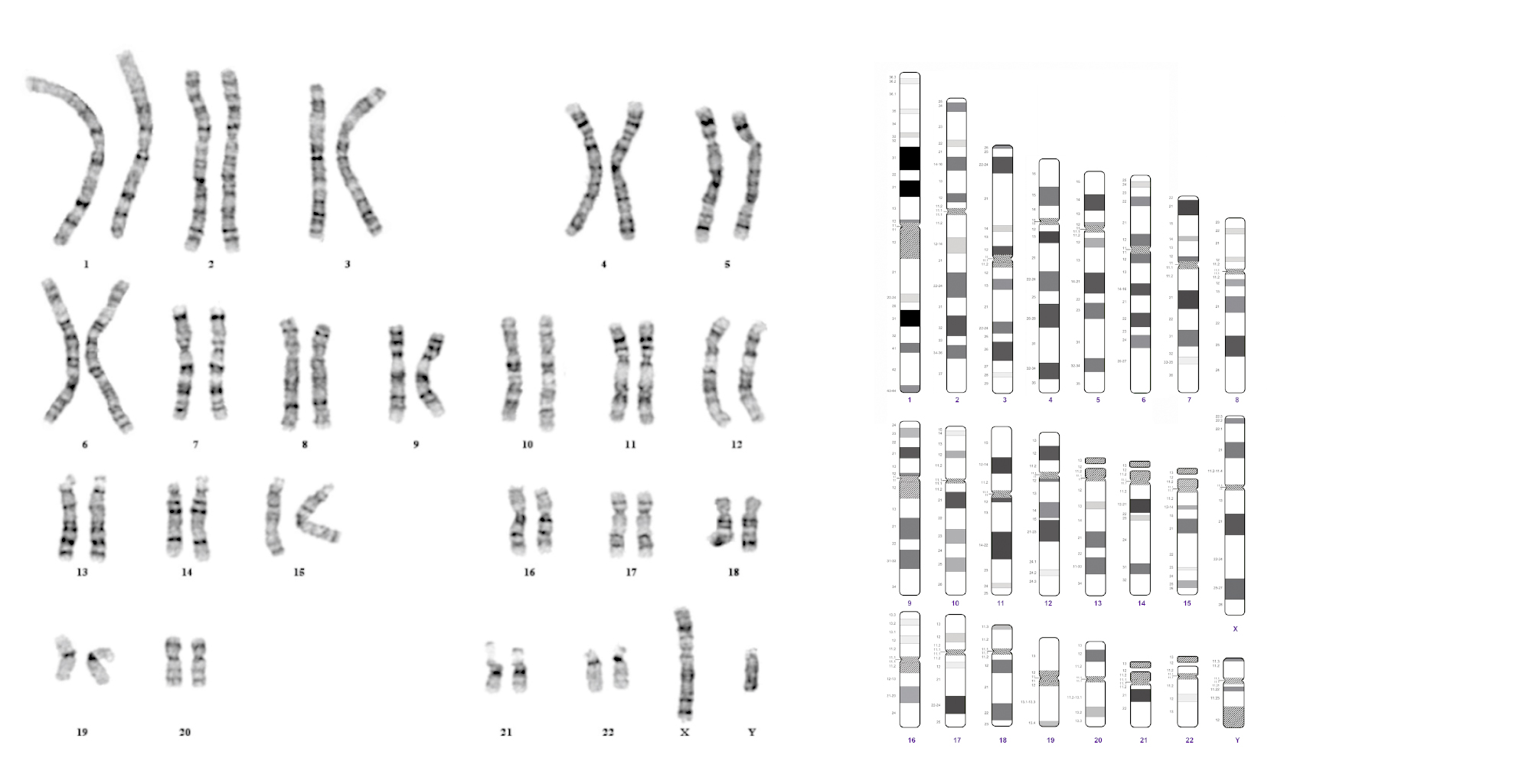
A citogenetika a kromoszómákkal, azok öröklődésben és betegségekben betöltött szerepével foglalkozó tudomány. A hagyományos vagy konvencionális citogenetika a kariotípus meghatározását jelenti. A kariotípus 20-25 metafázis fénymikroszkópos és számítógépes szoftverrel történő értékelésén alapszik. A kromoszómák preparálása Nátrium-heparinátos perifériás vér 72 órás limfocita tenyészetéből történik. Az osztódásához szükséges blasztos transzformációt a phytohemagglutinin nevű mitogénnel idézzük elő, majd az osztódást a sejtciklus metafázisban colchicinnel leállítjuk. Az osztódó sejtek hipotóniás oldattal kezelve megduzzadnak, maghártyájuk felszakad, és a kromoszómák elkülönülnek egymástól. Az egy sejthez tartozó kromoszómák együttmaradásához a sejtszuszpenziót metanol-ecetsav keverékével fixáljuk és többször átmossuk. A szuszpenziót tárgylemezre cseppentjük, pár napig szárítjuk, majd megsávozzuk. A legnépszerűbb sávozási eljárás az ún. Giemsa sávozás (G-sáv), mely során sóoldatos-tripszines kezeléssel a fehérjét „kimarjuk” a DNS-ből, a Giemsa-festék az adenin-timin nukleotidokban gazdag régiókban erősebb, a guanin-citozin nukleotidokban gazdag részekben halványabb jellegzetes „mintát”, változó sávozottságot hoz létre az egyes kromoszómákon. A kromoszómák átlagos felbontása 400-500 sáv (de elérheti akár a 850-et is), amely az összes számbeli és kb. 3-5 Mb nagyságrendű és feletti szerkezeti és számbeli kromoszóma rendellenesség azonosítására alkalmas. A veleszületett számbeli eltérések (teljes kromoszóma többlete vagy hiánya) gyakrabban érintik a nemi kromoszómákat (Turner-, Klinefelter szindróma) és ritkábban az autoszómákat (Down-, Patau- , Edwards-szindróma). A szerkezeti aberrációk lehetnek kiegyensúlyozottak (genomvesztés vagy többlet nélküli átrendeződések) és aneuploidiával járó kiegyensúlyozatlanok. A szerkezeti kromoszóma-rendellenességek az újszülöttek közel 1%-át érintik. Bizonyos rosszindulatú tumoros megbetegedésekben is jellegzetes kromoszóma szerkezeti vagy számbeli eltérés azonosítható.